Přibližné řešení úlohy Biochemické modely (10), Biochemické modely (11)
Charakteristickým rysem reakcí enzymu se substrátem je to, že koncentrace enzymu je výrazně menší, než koncentrace substrátu, To vzhledem k Biochemické modely (9) znamená, že
parametr je „skoro nula“. Také můžeme říci, že pravá strana druhé rovnice systému Biochemické modely (10) je „skoro nekonečno“, nebo že pravá strana první rovnice tohoto systému je zanedbatelně malá ve srovnání s pravou stranou druhé rovnice. Veličina
se mění „nesrovnatelně pomaleji“, než veličina
takže veličina
je vzhledem k
„skoro konstantní“. Z těchto důvodů budeme
ve druhé z rovnic systému Biochemické modely (10) považovat za konstantní parametr a tuto rovnici vyřešíme. Jedná se o rovnici se separovanými proměnnými, takže její řešení splňující druhou podmínku z dvojice Biochemické modely (11), tj. podmínku
dostaneme ve tvaru
|
|
(12) |
Platí pro ně
„Rychle se měnící“ proměnná veličina (funkce) se tedy „velice rychle“ ustálí na hodnotě
Ovšem hodnota
se také mění, i když „pomalu“. Tato změna je popsána první rovnicí systému Biochemické modely (10). V ní můžeme proměnou
považovat za parametr rovný ustálené hodnotě
S využitím počáteční podmínky Biochemické modely (11) tak dostaneme počáteční úlohu
Řešení této úlohy, které je „trochu jiné“ než řešení původní úlohy Biochemické modely (10), Biochemické modely (11) a proto ho označíme symbolem je implicitně dáno rovností
|
|
(13) |
Takto definovanou funkci můžeme považovat za první složku přibližného řešení úlohy Biochemické modely (10), Biochemické modely (11). Její druhou složku vyjádříme jako
|
|
(14) |
tato funkce však nesplňuje druhou z počátečních podmínek Biochemické modely (11).
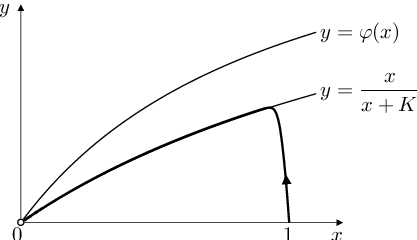 |
|
Obr. 5. Nulkliny systému Biochemické modely (10) a jeho trajektorie s počáteční podmínkou Biochemické modely (11) a hodnotou parametru
|
Funkce definované vztahy Biochemické modely (13) a Biochemické modely (14) se nazývá pseudo- nebo quasi-stacionární aproximace řešení úlohy Biochemické modely (10), Biochemické modely (11). V mnoha aplikacích je tato aproximace dostatečně přesná. Na obrázku Biochemické modely 5 je trajektorie řešení úlohy Biochemické modely (10), Biochemické modely (11) s hodnotou parametru
vidíme, že trajektorie řešení s „malou“ hodnotou parametru
skutečně od jistého bodu téměř splývá s
-nulklinou, tj. s funkcí
Řešení úlohy Biochemické modely (10), Biochemické modely (11) si tedy lze představit tak, že v „kratičkém časovém intervalu“ od začátku reakce se veličina (relativní množství substrátu) „nestačí změnit“, takže má stále počáteční hodnotu 1. V tomto „kratičkém čase“ veličina
(relativní množství komplexu
vzhledem k množství enzymu) rychle dosáhne své quasi-stacionární hodnoty. Tento „rychlý nárůst“ je popsán rovností Biochemické modely (12) do níž je dosazeno
quasi-stacionární hodnota je tedy
Dále se veličiny
a
vyvíjejí tak, jak je popsáno rovnostmi Biochemické modely (13) a Biochemické modely (14).
Ještě můžeme specifikovat délku zmíněného „kratičkého časového intervalu“ pro dosažení quasi-stacionárního stavu. Předpokládejme, že jsme schopni měřit koncentrace s relativní přesností Pak čas
za nějž veličina
naroste do quasi-stacionární hodnoty
je přibližně dána přibližnou rovnicí
tedy
|
|
(15) |
Popsanou aproximaci řešení lze získat i jiným způsobem méně se odvolávajícím na intuici: řešení úlohy Biochemické modely (10), Biochemické modely (11) budeme hledat ve tvaru Taylorových řad v proměnné Předpokládejme tedy, že řešení úlohy Biochemické modely (10), Biochemické modely (11) je tvaru
Za předpokladu, že tyto řady, chápané jako řady funkcí proměnné konvergují stejnoměrně (k tomu při
stačí, aby všechny funkce
byly ohraničené stejnou konstantou), platí
|
|
a současně
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Porovnáním koeficientů u stejných mocnin získáme nekonečný systém rovnic
Z první dvojice rovnic dostaneme
tedy quasi-stacionární aproximaci řešení Biochemické modely (13), Biochemické modely (14). V tomto případě však tato aproximace závisí na jedné integrační konstantě a ta závisí na počátečních podmínkách. Počáteční podmínky Biochemické modely (11) lze zapsat ve tvaru
takže z věty o jednoznačnosti Taylorových řad plyne
První z těchto podmínek lze splnit volbou stejně jako v Biochemické modely (13), ale druhou z nich splnit nelze. Odtud plyne, že alespoň jedna ze složek
řešení úlohy Biochemické modely (10), Biochemické modely (11) nemůže být analytickou funkcí parametru
Aby bylo možné splnit počáteční podmínky, je třeba v pravém okolí bodu hledat řešení úlohy Biochemické modely (10), Biochemické modely (11) jiným způsobem. Zavedeme novou nezávisle proměnnou
|
|
(16) |
Pro je
takže změnou časového měřítka Biochemické modely (16) „natáhneme malé okolí“
na „velice dlouhou dobu“. Substitucí Biochemické modely (16) se úloha Biochemické modely (10), Biochemické modely (11) transformuje na úlohu
|
|
(17) |
|
|
(18) |
Kvalitativní analýza úlohy Biochemické modely (17), Biochemické modely (11) dá stejné výsledky jako v ??.
Řešení úlohy Biochemické modely (17), Biochemické modely (18) budeme opět hledat ve tvaru Taylorových řad v proměnné tj. ve tvaru
Pak je
a současně
Z počátečních podmínek Biochemické modely (18) dostaneme
Nulté aproximace
řešení úlohy Biochemické modely (17), Biochemické modely (11) jsou řešením počáteční úlohy
takže
Vrátíme se k původní nezávisle proměnné a dostaneme novou aproximaci řešení úlohy Biochemické modely (10), Biochemické modely (11) ve tvaru
|
|
(19) |
tyto funkce splňují počáteční podmínky Biochemické modely (11).
Řešení úlohy Biochemické modely (10), Biochemické modely (11) lze v okolí bodu tj. na intervalu
pro vhodné malé kladné číslo
aproximovat funkcemi Biochemické modely (19). Tato část řešení úlohy se nazývá singulární nebo vnitřní řešení. Na intervalu
lze použít quasi-stacionární aproximaci Biochemické modely (13), Biochemické modely (14); tato část řešení úlohy se nazývá nesingulární nebo vnější řešení.
Na obr. Biochemické modely 6 je znázorněno přibližné a přesné řešení úlohy Biochemické modely (10), Biochemické modely (11) s parametrem vidíme, že již v tomto případě je přibližné řešení dosti blízké přesnému. Navíc, první složka řešení, tj. funkce
je i v pravém okolí nuly přesněji aproximována vnějším řešením než vitřním.
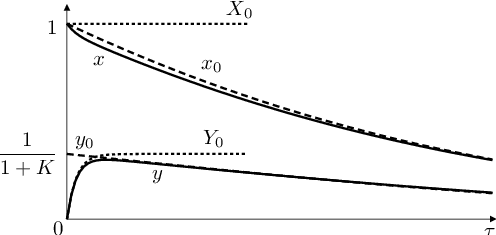 |
|
Obr. 6. Řešení úlohy Biochemické modely (10), Biochemické modely (11) s parametrem
|
Ještě odhadneme parametr - časový okamžik, od něhož vnější řešení lépe než vnitřní aproximuje druhou složku řešení úlohy Biochemické modely (10), Biochemické modely (11). Je to taková hodnota nezávisle proměnné, v níž mají funkce
a
stejnou hodnotu,
Takové číslo
existuje podle Bolzanovy věty, neboť
Můžeme tedy řešit soustavu rovnic
To znamená, že řešení druhé z rovnic, tj. rovnice leží v intervalu
a funkce
je na tomto intervalu rostoucí. Odtud dále plyne, že existuje konstanta
taková, že pro řešení
druhé z rovnic platí
Z první rovnice nyní dostaneme
Tato nerovnost vyjadřuje, že hodnota je malá stejného řádu, jako
tj.
Tento odhad souhlasí s vyjádřením Biochemické modely (15).
Řešení původní úlohy Biochemické modely (6), Biochemické modely (7) můžeme nyní zapsat ve tvaru
|
|
|
|
přitom funkce je implicitně dána rovnicí Biochemické modely (13).
