Základní modely enzymatické kinetiky
Uvažujme reakci nějakého substrátu a enzymu
které spolu vytvoří nestabilní komplex
z kterého dále vznikne nějaký produkt
a volný enzym. Enzymy
jsou katalyzátory chemických reakcí, při kterých pomáhají ze substrátu
vytvořit produkt
přičemž z reakce vycházejí samy v nezměněné formě.
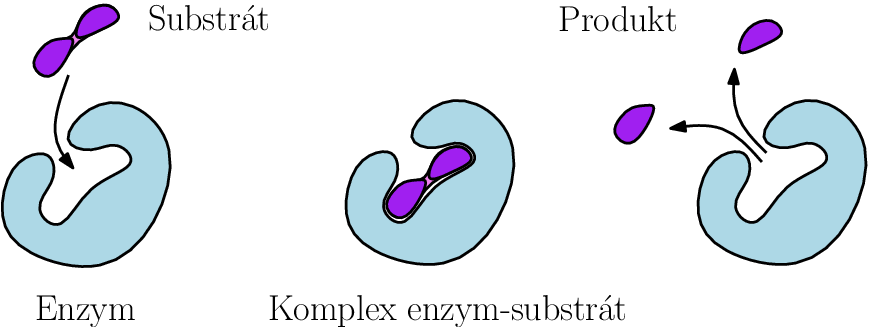
Schematicky tuto reakci můžeme zapsat takto
nebo stručně
přičemž kladné parametry
označují reakční rychlosti. První část reakce (vratná reakce tvorby komplexu) byla studována v předchozí kapitole.
Označme
Michaelis a Menten1 navrhli jako model vývoje koncentrací v čase následující systém čtyř obyčejných nelineárních diferenciálních rovnic
|
|
|
(2) |
|
|
|
|
|
|
|
|
|
|
|
Tento model vyjadřuje, že změny koncentrací považujeme za přímo úměrné koncentracím, reakční rychlosti jsou příslušné koeficienty úměrnosti. Budeme předpokládat, že na počátku je koncentrace substrátu rovna
a koncentrace enzymu je rovna
komplex
ani produkt
nejsou na počátku přítomny. Spolu se systémem Biochemické modely (2) tedy uvažujeme počáteční podmínky
|
|
(3) |
Nejprve si všimněme, že veličina se nevyskytuje v prvních třech rovnicích systému Biochemické modely (2). Koncentrace
a
jsou tedy řešením prvních tří rovnic z Biochemické modely (2), koncentraci produktu můžeme vyjádřit ze čtvrté rovnice integrací
|
|
(4) |
Množství enzymu se v průběhu reakce nemění a enzym se vyskytuje jednak jako volný a jednak jako vázaný v komplexu
To vzhledem k počáteční podmínce Biochemické modely (3) znamená, že by mělo platit
pro všechna
Model Biochemické modely (2) je skutečně v tomto smyslu adekvátní, neboť
Veličina je prvním integrálem systému Biochemické modely (2) a proto koncentraci enzymu můžeme vyjádřit jako
|
|
(5) |
a dosadit do první a třetí rovnice systému Biochemické modely (2). Dostaneme
|
|
(6) |
Časový průběh koncentrací substrátu a komplexu
je tedy řešením systému dvou obyčejných autonomních nelineárních diferenciálních rovnic Biochemické modely (6) s počáteční podmínkou
|
|
(7) |
průběh koncentrací volného enzymu a produktu
je dána výrazy Biochemické modely (5) a Biochemické modely (4).
Změníme měřítko tak, aby všechny veličiny byly bezrozměrné, tj. zavedeme substituci
|
|
(8) |
veličina vyjadřuje koncentraci substrátu a veličina
koncentraci komplexu
v jednotkách počáteční koncentrace substrátu a enzymu. Časová jednotka je určena rychlostí reakce substrátu a enzymu. Platí
|
|
|
|
|
|
|
|
Při označení
|
|
(9) |
se systém Biochemické modely (6) substitucí Biochemické modely (8) transformuje na systém
|
|
(10) |
s počátečními podmínkami
|
|
(11) |
Poznamenejme, že parametry
a
jsou kladné a
Úlohu Biochemické modely (10), Biochemické modely (11) nelze řešit explicitně. Proto ji budeme analyzovat ve fázovém prostoru. Nulklinu proměnné můžeme vyjádřit jako graf funkce
|
Derivace
je pro
|
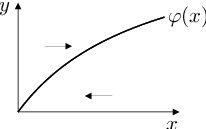 |
|
Podobně vyjádříme a vyšetříme znaménka derivací:
|
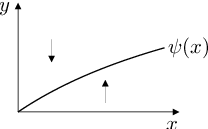 |
Poněvadž a
pro všechna
vypadá fázový portrét systému Biochemické modely (10) tak, jak je znázorněno na obr. Biochemické modely 3 Vidíme, že systém Biochemické modely (10) má jediný stacionární bod
a že množina
je jeho pozitivně invariantní množinou (na úsečce
směřují trajektorie nahoru, na úsečce
dolů, na úsečce
směřují trajektorie doprava a na úsečce
doleva). Variační matice systému Biochemické modely (10) v obecném bodě
je
takže ve stacionárním bodě platí
|
|
|
|
|
|
|
|
|
neboť Stacionární bod
je podle Kvalitativní vlastnosti řešení dvourozměrného autonomního systému Autonomní systémy a kvalitativní teorie (5) stabilní uzel (což bylo vidět z fázového portrétu i bez výpočtů). Pro řešení úlohy Biochemické modely (10), Biochemické modely (11) tedy platí
Výsledkem reakce je vyčerpání veškerého substrátu nebude volný ani vázaný s enzymem v komplexu
Z trajektorie řešení úlohy Biochemické modely (10), Biochemické modely (11), která je rovněž zobrazena na následujícím obrázku, je také vidět, že složka
řešení této úlohy k nule monotonně klesá. Složka
nejdříve roste, v jistém čase
dosáhne svého maxima
a pak monotonně klesá k nule.
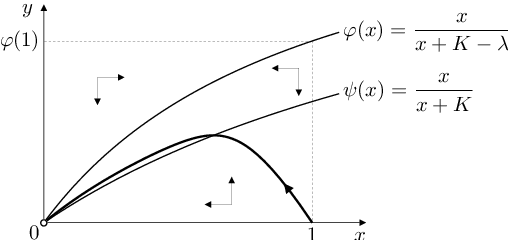 |
|
Obr. 3. Fázový portrét systému Biochemické modely (10) a jeho trajektorie s počátečním bodem Biochemické modely (10) a hodnotou parametru
|
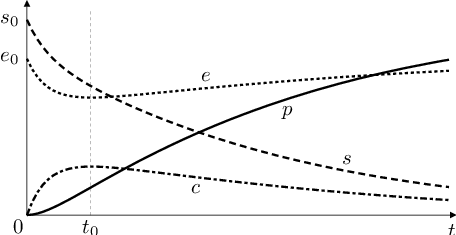
Nyní můžeme kvalitativně popsat řešení původní úlohy Biochemické modely (2), Biochemické modely (3), viz obr. následující obrázek. Koncentrace substrátu
monotonně klesá k nule. Koncentrace
komplexu
roste ke své maximální hodnotě, která je menší než byla počáteční koncentrace
enzymu
a pak monotonně klesá k nule. Koncentrace
volného enzymu
nejprve klesá, v okamžiku
kdy je koncentrace komplexu
maximální, dosáhne svého minima a pak monotonně roste k počáteční hodnotě
Koncentrace
produktu
roste z nulové hodnoty, růst se nejprve zrychluje (funkce je konvexní), od okamžiku
se začne zpomalovat (funkce je konkávní).
 |
|
Obr. 4. Průběh řešení úlohy Biochemické modely (2),Biochemické modely (3).
|
1 L.Michaelis, M. I. Menten. Die Kinetik der Invertinwirkung. Biochem. Z. 49, 333-369, 1913
